The Pathobyte Series: Francisella tularensis: Secret Life of the Ultra-Infectious Living Corkscrew
Francisella tularensis, an ultra-infectious, pleomorphic bacterium, historically mimicked ancient plagues and confused early doctors before being isolated by Edward Francis. As a fastidious mesophile, it resiliently survives environmental freezing inside biofilms but strictly requires host cysteine to successfully replicate. Inside the human body, it turns macrophages into private nurseries, deploying a Type VI secretion system to escape destruction and cause deadly tularemia. Its unique genetic vulnerabilities inspired historical live attenuated vaccines and modern rapid NAAT diagnostics. Though recognized as a potent respiratory biothreat famously aerosolized by active lawn mowers, it completely lacks any documented human-to-human transmission pathways.
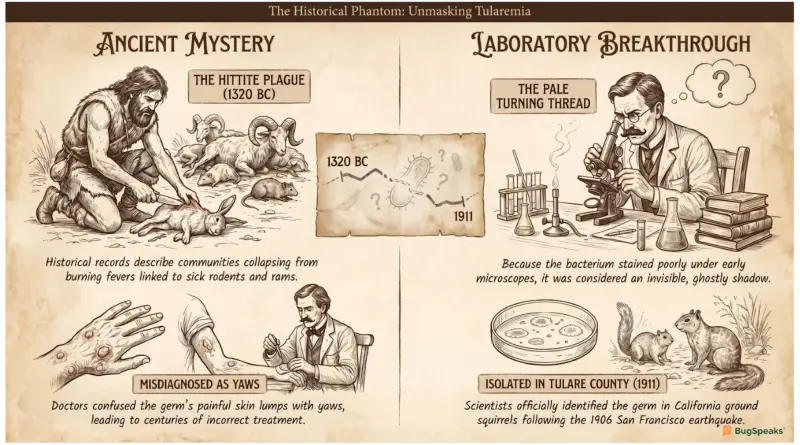
Which Ghostly Shadow Bluffed Doctors and Mimicked Ancient Tropical Plagues?
Long before scientists had high-tech microscopes, ancient populations suffered from a mysterious, invisible threat that modern historians believe was caused by a dangerous pathogen. This sneaky microorganism caused a terrifying sickness that swept through armies and villages, showing up unexpectedly in wild animal populations. In ancient Central Anatolia, historical records describe a devastating event known today as the "Hittite Plague," where communities collapsed from a strange, burning fever. People noticed that when local wild animals like rams and rodents got sick, humans would quickly fall ill too, though nobody understood how this tiny enemy travelled.
Centuries later in Europe, the same invisible killer re-emerged, frustrating doctors because it was completely invisible under early, basic magnifying tools. Because the germ stained very poorly and looked so faint, it was called a pale, ghostly shadow that couldn't be caught. It was frequently confused with a completely different tropical disease called yaws because both illnesses caused painful, open skin lumps and swollen bodies. This confusion meant that for generations, people treated the illness completely incorrectly, allowing the phantom bacterium to spread quietly through families, hunters, and farmers without ever being truly identified.
The true breakthrough came during routine animal health surveillance following the massive 1906 San Francisco earthquake, when researchers noticed ground squirrels dying by the thousands. Scientists named the microscopic culprit after Tulare County, California, where they officially isolated it, finally proving it wasn't the famous black plague germ. A brave bacteriologist named Edward Francis dedicated his entire life to mapping this illness, which we now call tularemia in his honor. Amazingly, the germ was so incredibly infectious that Dr. Francis and every single one of his laboratory assistants accidentally caught it while investigating its secrets.
How Does a Broken Fuel Engine Force This Organism to Beg for Host Micro-Nutrients?
Even though this germ is an obligate parasite that must cause disease to thrive, it displays a truly stunning survival strategy outside its host. In nature, it occupies a massive geographic range across the Northern Hemisphere, hiding inside more than two hundred different animal species. It acts as a mesophilic creature, meaning it grows best at warm, cozy body temperatures between 35°C and 37°C. However, when it finds itself trapped outside a warm body, it doesn't just wither away; instead, it enters a deep, protective sleep state to survive environmental stress.
When facing freezing cold waters, the germ forms a slimy, defensive neighborhood called a biofilm or turns into a special "viable but non-culturable" state. This means the bacterium stays alive but stops growing, surviving for years in frozen meat or chilly rivers by hiding inside free-living amoebae. To stay safe from predators, it forms partnerships with these tiny water creatures, which shield the germ from harsh chemicals and water treatment plants.
Because the bacterium has a broken internal system, it has an absolute, non-negotiable requirement for an external nutrient called cysteine to build proteins. It cannot make this amino acid on its own, so it must steal it directly from the liquid inside a host cell. Without a living host to provide this constant fuel, the fragile microbe would eventually starve and lose its ability to multiply. This intense dependence on host nutrients is exactly why it has evolved such aggressive, high-speed methods to break inside our cells and hijack our internal supplies.
What Secret Weapon Allows a Shape-Shifting Phantom to Trick and Explode Human Immune Cells?
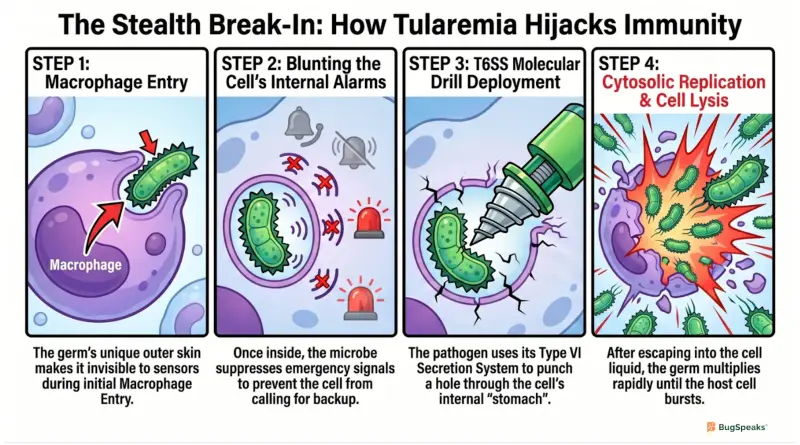
The shape of this microbe is pleomorphic, which means it can morph its tiny body into a circle or a short rod. It acts as an aerobic organism that needs oxygen to burn its fuel, but it acts like a secret double agent inside us. Once it enters the bloodstream, it seeks out big immune cells called macrophages, which are supposed to eat and destroy invading germs. Instead of running away, this tiny living corkscrew willingly lets itself get swallowed up, turning our body's defense shields into its own private, cozy nursery.
To pull off this amazing trick, the germ uses a specialized weapon called a capsule along with an incredibly weird outer coat. This unique outer skin is completely invisible to our body's warning sensors, meaning it never triggers the emergency alarms that call for backup. Once inside the immune cell's stomach, the germ uses a tiny molecular drill called a Type VI secretion system to punch a hole right through the stomach wall. To see exactly how this sneaky break-in happens step-by-step, follow the interactive pathway below:
[Germ gets eaten] ──► [Blunts cell alarms] ──► [Drills out of stomach] ──► [Multiplies in cell liquid]
Once it escapes into the cell's main liquid chamber, it uses a special enzyme to feast on the host's energy supplies. It multiplies rapidly until the cell gets stuffed to maximum capacity, eventually bursting open and sending millions of new germs into the body. This destruction leads to a severe condition called lymphadenopathy, where the body's lymph nodes swell up like painful, solid marbles. If the microbes are inhaled through dust, they cause a deadly lung infection that can easily end a life if left completely untreated.
Can Cooking a Defenseless Germ from the Inside Out Reveal the Vulnerabilities of Dangerous Pathogens?
Long before modern antibiotics existed, doctors discovered a bizarre trick to treat this infection: they intentionally gave patients a temporary, blazing fever. This worked because the germ has a major genetic flaw; it completely lacks heat-shock regulators, which are emergency shield proteins that other bacteria use to survive high heat. When a patient's body temperature soared, the defenseless bacteria would literally cook and fall apart inside the blood. This discovery proved that studying the weaknesses of a single fastidious microbe could teach scientists how to manipulate human immune responses to fight off deep infections.
Later, scientists used this germ to develop early versions of an attenuated vaccine, which uses a weakened live strain to train the body. By intentionally deleting two specific regions of the germ's DNA, researchers created a safe version that can't cause severe illness. This medical milestone helped launch the historical era of "magic bullets," showing that doctors could use tailored medicine to target specific bacterial parts without harming the patient.
Today, scientists use these historical lessons to build incredibly precise medical tools that keep laboratory workers and communities completely safe from outbreaks. By extracting the germ's unique outer coat using hot water methods, researchers can create custom antibodies that act like heat-seeking missiles. These engineered molecules are packed into rapid test strips that can instantly spot the pathogen in mysterious powders or dirty water. This old battle against a tiny corkscrew completely reshaped modern biotechnology, turning a dangerous threat into a teacher for human survival.
Can We Turn a Pathogen's Unique DNA Sequence Into an Instant Warning Alarm?
For over a hundred years, scientists struggled to grow this fastidious organism inside laboratories because it refused to multiply on standard petri dishes. This happened because the microbe has a strictly microaerophilic requirement, meaning it suffocates if there is too much or too little oxygen around. To solve this mystery, modern geneticists mapped out the germ's entire DNA blueprint, discovering a goldmine of information hidden inside its code. They found that the bacterium relies on a specialized enzyme called gamma-glutamyl transpeptidase to steal nutrients directly from the cells of living rabbits.
By understanding this genetic blueprint, scientists invented advanced diagnostic tests called NAAT methods, which stand for nucleic acid amplification tests. These high-tech tools scan a sample and replicate a tiny piece of the germ's unique DNA millions of times over. Instead of waiting weeks for a slow microbe to grow in a dish, doctors can now identify the infection in just a few minutes. This rapid detection allows hospitals to start life-saving treatments immediately, long before the sneaky living corkscrew has a chance to drill deep into our internal organs.
These genetic maps are also helping international researchers design a permanent, high-tech shield that could wipe out this disease forever. By studying the exact shape of the proteins on the germ's outer skin, scientists are engineering a modern, synthetic vaccine. This ultimate shield will teach our white blood cells to recognize and destroy the living corkscrew the absolute millisecond it enters our body. Through teamwork, curiosity, and genetic science, humanity is turning the tide against this ancient warrior, ensuring that the invisible thread will never terrify the world again.
Taxonomic Classification Table
Microbe Profile
Shape: Extremely small, pleomorphic coccobacillus or short rod
Gram Stain Nature: Gram-negative, staining very faintly and poorly with standard laboratory dyes.
Spore-forming: Non-spore-forming.
Biofilm formation: Fully capable of forming functional biofilms to survive long-term in freshwater reservoirs and plumbing systems.
Oxygen requirements: Aerobic respiration metabolism, but operates as a facultative intracellular pathogen inside host cells.
Optimal Temperature: Mesophilic, growing optimally between 35°C and 37°C.
Optimal pH: Neutrophilic, with optimal growth between pH 6.8 and 7.4; possesses robust acid-resistance genes for low-pH survival.
Nutrient Usage: Highly fastidious; has a strict requirement for external cysteine due to a broken sulfate pathway caused by an adenylylsulfate kinase pseudogene. It uses host cytosolic glutathione via gamma-glutamyl transpeptidase during replication. It is oxidase-negative, weakly catalase-positive, urease-negative, lacks a functional pfkA gene, and uses the Embden-Meyerhof-Parnas pathway for gluconeogenesis.
Fun Facts
The Lawn Mower Menace: In areas where this microbe lives, simply mowing the lawn is a recognized medical risk factor! Running over an infected rodent carcass can turn the germ into an invisible mist, which people accidentally breathe in, leading directly to a deadly lung infection.
The Ultra-Resilient Freeze Master: This pathogen is unbelievably tough when things get chilly. Scientists discovered that it can stay perfectly alive and ready to infect inside frozen rabbit meat for more than three whole years!
The Lonely Attacker: Despite being one of the most infectious and dangerous respiratory biothreat agents on the planet, human-to-human transmission has never been documented. You can only catch it from nature, animals, or vectors, never from another person's cough.
Reference
Di Spirito, M., Pascolini, C., Salemi, S., Spagnolo, F., Luca, V., Molinari, F., Rozov, O., Lista, F., D’Amelio, R., & Fillo, S. (2026). Tularemia: Historical Perspectives and Current Challenges of a Re-Emerging Zoonosis. Biomedicines, 14(3), 695. https://doi.org/10.3390/biomedicines14030695
Degabriel, M., Valeva, S., Boisset, S., & Henry, T. (2023). Pathogenicity and virulence of Francisella tularensis. Virulence, 14(1), 2274638. https://doi.org/10.1080/21505594.2023.2274638
Gürcan S. (2014). Epidemiology of tularemia. Balkan medical journal, 31(1), 3–10. https://doi.org/10.5152/balkanmedj.2014.13117
Männikkö N. (2011). Etymologia: Francisella tularensis. Emerging Infectious Diseases, 17(5), 799. https://doi.org/10.3201/eid1705.ET1705
Hirschmann J. V. (2018). From Squirrels to Biological Weapons: The Early History of Tularemia. The American journal of the medical sciences, 356(4), 319–328. https://doi.org/10.1016/j.amjms.2018.06.006
Freudenberger Catanzaro, K. C., & Inzana, T. J. (2020). The Francisella tularensis Polysaccharides: What Is the Real Capsule?. Microbiology and molecular biology reviews : MMBR, 84(1), e00065-19. https://doi.org/10.1128/MMBR.00065-19
McLendon, M. K., Apicella, M. A., & Allen, L. A. (2006). Francisella tularensis: taxonomy, genetics, and Immunopathogenesis of a potential agent of biowarfare. Annual review of microbiology, 60, 167–185. https://doi.org/10.1146/annurev.micro.60.080805.142126
Sharma, R., Patil, R. D., Singh, B., Chakraborty, S., Chandran, D., Dhama, K., Gopinath, D., Jairath, G., Rialch, A., Mal, G., Singh, P., Chaicumpa, W., & Saikumar, G. (2023). Tularemia - a re-emerging disease with growing concern. The veterinary quarterly, 43(1), 1–16. https://doi.org/10.1080/01652176.2023.2277753
Zhang, X. H., Ahmad, W., Zhu, X. Y., Chen, J., & Austin, B. (2020). Viable but nonculturable bacteria and their resuscitation: implications for cultivating uncultured marine microorganisms. Marine life science & technology, 3(2), 189–203. https://doi.org/10.1007/s42995-020-00041-3
Pohanka, M. (2026). Immunosensors and Immunoassays to Detect Francisella tularensis and Diagnose Tularemia. Biosensors, 16(3), 158. https://doi.org/10.3390/bios16030158
Lu, Z., Rynkiewicz, M. J., Yang, C. Y., Madico, G., Perkins, H. M., Roche, M. I., Seaton, B. A., & Sharon, J. (2014). Functional and structural characterization of Francisella tularensis O-antigen antibodies at the low end of antigen reactivity. Monoclonal antibodies in immunodiagnosis and immunotherapy, 33(4), 235–245. https://doi.org/10.1089/mab.2014.0022
Meibom, K. L., & Charbit, A. (2010). Francisella tularensis metabolism and its relation to virulence. Frontiers in microbiology, 1, 140. https://doi.org/10.3389/fmicb.2010.00140
Meibom, K. L., & Charbit, A. (2010). Francisella tularensis metabolism and its relation to virulence. Frontiers in microbiology, 1, 140. https://doi.org/10.3389/fmicb.2010.00140
Coolen, J. P., Sjödin, A., Maraha, B., Hajer, G. F., Forsman, M., Verspui, E., Frenay, H. M., Notermans, D. W., de Vries, M. C., Reubsaet, F. A., Paauw, A., & Roeselers, G. (2013). Draft genome sequence of Francisella tularensis subsp. holarctica BD11-00177. Standards in genomic sciences, 8(3), 539–547. https://doi.org/10.4056/sigs.4217923