Exploring the Connection Between Skinny Bacteria and Weight Loss
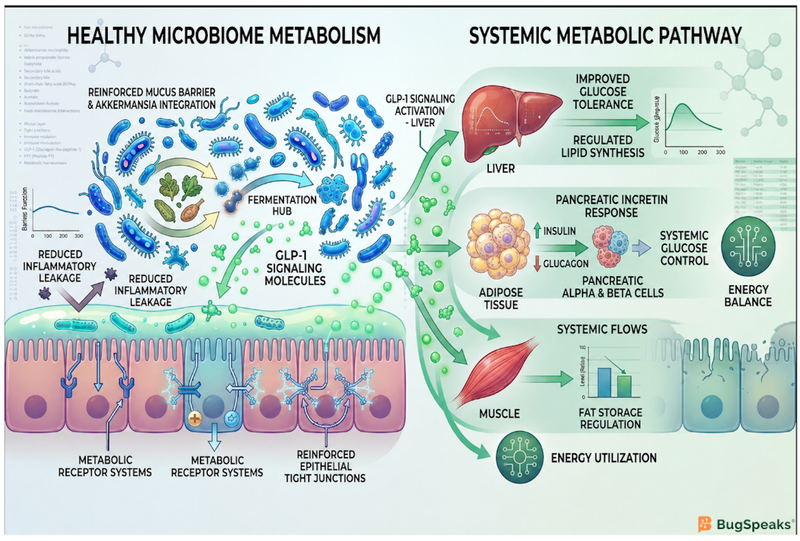
How Do Gut Bacteria Control Your Daily Energy Balance?
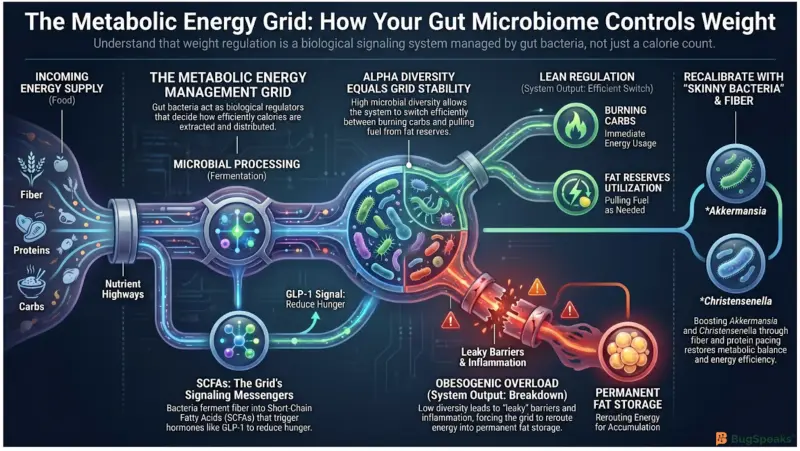
Gut bacteria control your daily energy balance by acting as regulators inside a Metabolic Energy Management Grid that determines how much incoming food is burned for fuel versus stored as fat. In this system, the food you eat becomes the incoming energy supply, while the gut microbiome operates as a network of microbial regulators that decide how efficiently calories are extracted, processed, and distributed throughout the body. Depending on which bacteria dominate the grid, the exact same meal can either support balanced metabolism or overload the body’s energy storage reserves.
A major part of this regulation begins with dietary fiber. Humans cannot digest many complex fibers on their own, but specific gut microbes contain carbohydrate-active enzymes that ferment these fibers into Short-Chain Fatty Acids (SCFAs) such as acetate, propionate, and butyrate (Masi et al., 2026). These molecules can provide up to 10% of daily human energy needs while also functioning as metabolic signaling molecules that influence appetite, fat burning, and insulin sensitivity.
SCFAs help stabilize the city-wide energy distribution network by activating receptors on intestinal cells called Free Fatty Acid Receptor 2 and 3. This activation stimulates the release of important hormones including Glucagon-Like Peptide-1 (GLP-1) and Peptide YY (PYY), which communicate with the brain to reduce hunger and improve blood sugar regulation (Masi et al., 2026). When the grid functions efficiently, these signals help prevent excess calorie intake and reduce unnecessary fat storage.
The overall performance of the grid depends heavily on alpha diversity, which measures how many different microbial species exist in the gut and how evenly they are distributed. A highly diverse microbiome contains multiple specialized regulators capable of processing different nutrients without overwhelming the system. This flexibility allows the body to switch efficiently between burning carbohydrates for immediate energy and pulling fuel from fat reserves when food intake drops.
When microbial diversity collapses, the grid becomes unstable. SCFA production declines, the intestinal barrier weakens, and appetite-regulating signals become less effective (Masi et al., 2026). As a result, the body begins storing more incoming calories inside adipose tissue while simultaneously increasing hunger signals. Over time, the energy management grid shifts away from balanced fuel usage and toward chronic energy retention.
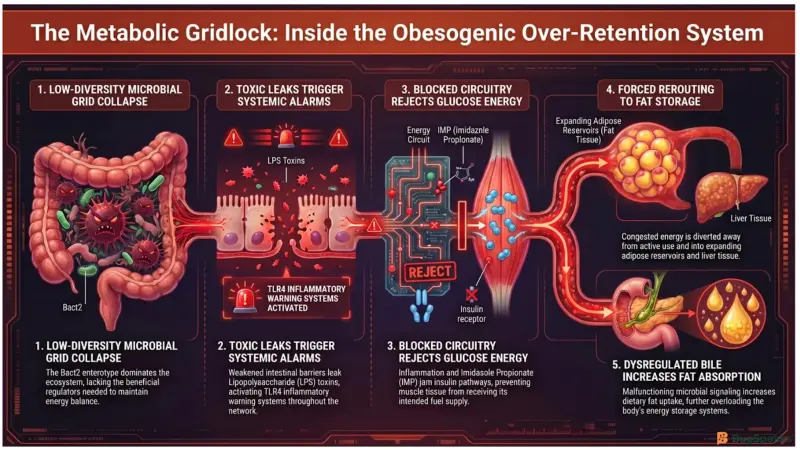
Why Do Obesogenic Microbes Store More Fat Than Needed?
Obesogenic microbes store more fat than needed because they function as over-retention systems that aggressively harvest calories while driving chronic inflammation inside the Metabolic Energy Management Grid. Instead of supporting balanced energy distribution, these microbial configurations push the body toward long-term energy storage and insulin resistance.
One of the clearest examples is the Bacteroides 2 (Bact2) enterotype, a low-diversity microbial pattern strongly associated with obesity and metabolic dysfunction (Masi et al., 2026). This configuration contains elevated levels of pro-inflammatory bacteria while lacking many beneficial SCFA-producing species. Without enough SCFA production, the intestinal barrier weakens and becomes more permeable.
This barrier breakdown allows bacterial toxins called Lipopolysaccharides (LPS) to leak into the bloodstream, a condition known as metabolic endotoxemia (Masi et al., 2026). Once inside circulation, LPS molecules activate Toll-Like Receptor 4 (TLR4) on human cells, triggering persistent low-grade inflammation throughout the body. In the energy grid, this inflammation acts like damage spreading through electrical infrastructure, disrupting how cells respond to insulin.
Because insulin controls how energy enters muscle cells, inflammation-driven insulin resistance prevents active tissues from efficiently using circulating glucose. The body responds by rerouting excess energy into adipose tissue, causing expansion of the energy storage reserves. At the same time, obesogenic microbes can alter bile acid metabolism, increasing dietary fat absorption and promoting additional fat accumulation in the liver and adipose tissue (Masi et al., 2026).
Certain obesogenic bacteria also produce metabolites such as Imidazole Propionate (IMP), which directly interfere with insulin signaling pathways in the liver (Masi et al., 2026). This further weakens metabolic control and reinforces the body’s tendency to retain energy rather than burn it. As long as these over-retention systems dominate the grid, the body remains locked in a persistent fat-storage state.
What Makes "Skinny Bacteria" Like Akkermansia So Effective at Weight Management?
“Skinny bacteria” like Akkermansia muciniphila support weight management by functioning as efficient balancing controllers that strengthen the intestinal barrier, reduce inflammation, and improve metabolic signaling throughout the energy grid. Rather than simply digesting food, these microbes actively maintain the structural integrity of the gut environment itself.
Akkermansia muciniphila lives within the intestinal mucus layer and feeds on mucin proteins rather than directly competing for dietary nutrients. Although it degrades mucus, this process stimulates intestinal cells to produce a thicker and healthier mucus barrier in return (Paone et al., 2026). In obesity-inducing high-fat diets, Akkermansia helps restore important mucus proteins such as Muc3, reinforcing the barrier that separates gut microbes from the bloodstream (Paone et al., 2026).
Strengthening this barrier prevents inflammatory leaks that would otherwise destabilize the energy grid. Akkermansia also produces an outer membrane protein called Amuc_1100, which activates Toll-Like Receptor 2 (TLR2) on intestinal cells (Huang et al., 2026). This signaling pathway tightens cellular junctions, reduces inflammation, and improves metabolic function. Remarkably, even pasteurized Akkermansia cells can preserve some of these benefits because the Amuc_1100 protein remains biologically active (Huang et al., 2026).
Beyond barrier repair, Akkermansia reshapes the chemical environment of the gut. It alters the activity of glycosyltransferases, enzymes responsible for attaching sugar molecules to intestinal mucus (Paone et al., 2026). These changes influence mucus sialylation and fucosylation patterns, helping beneficial microbes thrive while limiting the ability of harmful pathogens to attach to the intestinal wall. This creates a stable ecosystem that supports broader SCFA production across the microbial community.
Akkermansia also secretes a signaling protein called P9, which stimulates the release of GLP-1 and activates calorie-burning brown adipose tissue (Huang et al., 2026). Human trials additionally show that interventions enriching Akkermansia populations can improve intestinal barrier integrity in adults with obesity, further supporting its role as a major metabolic regulator (Batacan et al., 2026). By simultaneously suppressing appetite and increasing energy expenditure, these efficient balancing controllers help maintain a leaner metabolic profile while protecting the grid from inflammatory overload.
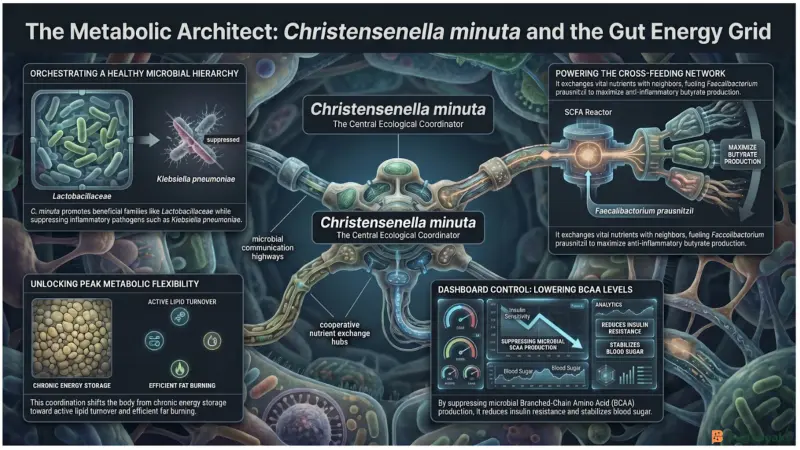
How Does Christensenella Minuta Act as a Master Energy Regulator?
Christensenella minuta acts as a master energy regulator by coordinating microbial cooperation throughout the Metabolic Energy Management Grid. Instead of operating independently, it functions as a central ecological organizer that improves communication, nutrient sharing, and metabolic efficiency across the gut ecosystem.
This bacterium is strongly associated with lower body weight and appears highly heritable in humans (Xu et al., 2024). Once established in the gut, Christensenella minuta reshapes the microbial community by promoting beneficial bacterial families such as Lactobacillaceae and Oscillospiraceae while suppressing opportunistic pathogens like Klebsiella pneumoniae (Xu et al., 2024). This stabilizes the grid and reduces inflammatory disruption.
Christensenella minuta also participates in extensive cross-feeding interactions. Because it cannot efficiently produce certain compounds such as vitamin B12 and serine, it depends on neighboring microbes for metabolic support (Xu et al., 2024). In return, it provides growth-promoting metabolites to other beneficial bacteria including Faecalibacterium prausnitzii, an important anti-inflammatory butyrate producer. These cooperative exchanges improve SCFA output and strengthen overall metabolic stability.
Another major effect involves the regulation of Branched-Chain Amino Acids (BCAAs). Elevated circulating BCAAs are strongly linked to insulin resistance and obesity risk (Masi et al., 2026). Christensenella minuta suppresses microbial pathways involved in excessive BCAA production, helping maintain insulin sensitivity and improving fat metabolism (Xu et al., 2024).
As the grid becomes more stable, the body shifts toward increased lipid turnover and beta-oxidation rather than chronic fat storage. Blood sugar regulation improves, circulating lipids decline, and the energy management system becomes more resistant to metabolic overload.
How Can You Reprogram Your Gut's Energy Grid Through Diet?
You can reprogram your gut’s energy grid through dietary strategies that selectively suppress obesogenic microbes while strengthening beneficial metabolic regulators. Among the most effective approaches studied so far are Intermittent Fasting (IF) and Protein Pacing (IF-P).
Clinical studies comparing continuous calorie restriction with IF-P show that intermittent fasting combined with structured protein intake produces greater reductions in visceral fat while preserving lean muscle mass (Mohr et al., 2024). These improvements are accompanied by major shifts in the gut microbiome, including increased populations of Christensenellaceae and Rikenellaceae, both associated with healthier lipid metabolism and reduced obesity risk.
During fasting periods, the temporary halt in incoming energy supply forces the body to mobilize stored fuel. This process increases signaling molecules such as Interleukin-4 (IL-4) and Interleukin-6 (IL-6), which help release fatty acids from adipose tissue for energy use (Mohr et al., 2024). At the same time, blood metabolites including acetylcarnitine rise, indicating enhanced fat oxidation and improved metabolic flexibility.
Protein pacing further stabilizes the energy grid by providing evenly distributed amino acid availability throughout the day. This supports lean muscle maintenance while also encouraging microbial pathways involved in protein and fat metabolism. Combined with increased fiber intake and reduced refined sugar consumption, the IF-P strategy weakens inflammatory over-retention systems and promotes a microbiome more favorable to metabolic health.
Long-term studies suggest these dietary shifts can produce durable restructuring of the microbial ecosystem. Even after substantial weight loss, individuals following IF-P protocols maintain increased populations of beneficial bacteria such as Akkermansia muciniphila and Anaerostipes hadrus for extended periods (Mohr et al., 2024). By controlling both nutrient timing and nutrient quality, the body gradually installs a more resilient and metabolically efficient energy management grid.
Visualize the process- https://youtu.be/IeI39gDliLw
Reference
Paone, P., Petitfils, C., Puel, A., Latousakis, D., de Vos, W. M., Delzenne, N. M., Juge, N., Van Hul, M., & Cani, P. D. (2026). Akkermansia muciniphilamodulates intestinal mucus composition to counteract high-fat diet-induced obesity in mice. Gut microbes, 18(1), 2612580. https://doi.org/10.1080/19490976.2025.2612580
Batacan, R., Rao, A., Bajagai, Y. S., Stanley, D., & Briskey, D. (2026). Oleoylethanolamide supplementation enriches Akkermansia muciniphila and modulates intestinal barrier function in adults with obesity: A randomized, double-blind, placebo-controlled trial. Gut Microbes Reports, 3(1). https://doi.org/10.1080/29933935.2026.2622259
Huang, B., Xian, Y., Xue, W., Pu, Z., Kong, P., Li, P., Zhao, Y., Tai, L., Chen, Z., Lan, Z., Liu, H. W., Jiang, X., & Chen, A. J. (2026). Probiotic attributes and safety profile of AKM Lab-01: a novel Akkermansia muciniphila strain combating obesity and metabolic disorders in diet-induced obese mice. Frontiers in microbiology, 16, 1627870. https://doi.org/10.3389/fmicb.2025.1627870
Mohr, A. E., Sweazea, K. L., Bowes, D. A., Jasbi, P., Whisner, C. M., Sears, D. D., Krajmalnik-Brown, R., Jin, Y., Gu, H., Klein-Seetharaman, J., Arciero, K. M., Gumpricht, E., & Arciero, P. J. (2024). Gut microbiome remodeling and metabolomic profile improves in response to protein pacing with intermittent fasting versus continuous caloric restriction. Nature communications, 15(1), 4155. https://doi.org/10.1038/s41467-024-48355-5
Xu C, Jiang H, Feng L-J, Jiang M-Z, Wang Y-L and Liu S-J (2024) Christensenella minuta interacts with multiple gut bacteria. Front. Microbiol. 15:1301073. doi: 10.3389/fmicb.2024.1301073
Masi, D., Watanabe, M., & Clément, K. (2026). Gut microbiome and obesity care: Bridging dietary, surgical, and pharmacological interventions. Cell reports. Medicine, 7(2), 102573. https://doi.org/10.1016/j.xcrm.2025.102573