The Ultimate Guide to Overcoming Constipation: Beyond Just Water
Why is water alone not enough to prevent constipation?
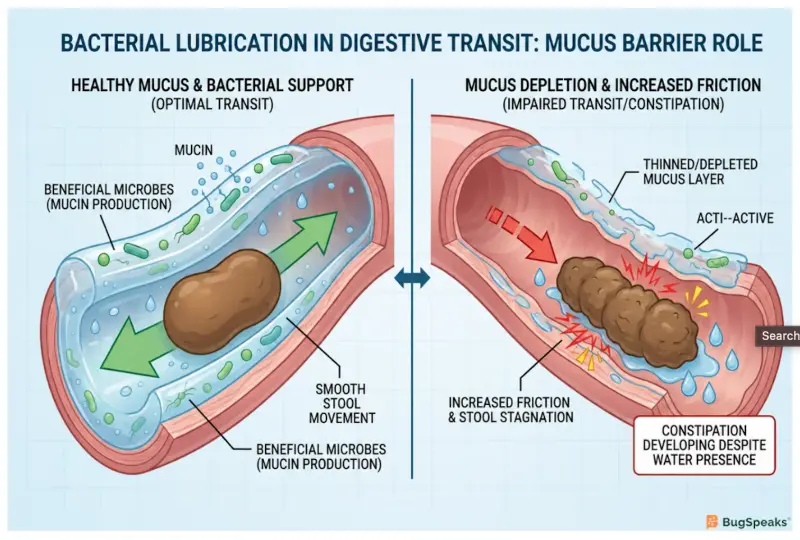
Water alone is insufficient because smooth intestinal movement also requires an intact mucosal barrier, which acts as a lubricating layer, and proper microbial balance to maintain this essential lubrication. To understand this biological reality, visualize the human colon as a highly regulated canal network. The solid waste, or transported material, cannot glide through this network based on water volume alone. The physical walls of the canal require a constant, protective natural flow coating known scientifically as mucin. Mucin is a glycoprotein that retains large amounts of water and shields the delicate epithelial cells from mechanical stress and dehydration (Hamaguchi et al., 2026). When we assume that reduced flow efficiency is merely a hydration problem, we ignore the biological necessity of this inner lining.
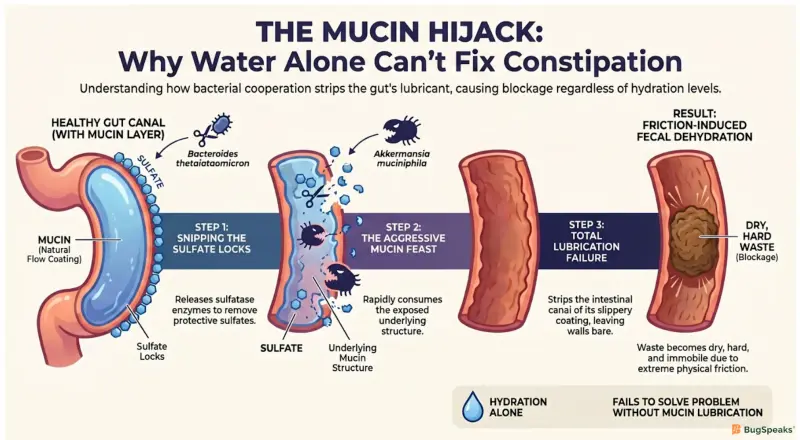
The health of this natural flow coating is entirely governed by our resident bacteria, which act as regulators of flow maintenance. Under healthy conditions, these microbes perfectly balance the creation and natural recycling of the mucosal gel. However, a specific biological malfunction occurs when two commensal bacteria, Akkermansia muciniphila and Bacteroides thetaiotaomicron, cooperatively over-degrade this protective layerHamaguchi et al. (2026). Colonic mucin features complex molecular structures capped with terminal sulfates. Because Akkermansia muciniphila lacks the specialized sulfatase enzymes required to detach these sulfates, it cannot consume colonic mucus entirely on its own.
The dynamic changes dangerously when Bacteroides thetaiotaomicron provides these missing sulfatase enzymes to the environment. Once the terminal sulfates are removed, Akkermansia muciniphila rapidly consumes the underlying mucin structure. This cooperative breakdown strips the canal network of its natural flow coating, leading directly to extreme fecal dehydration and severely reduced flow efficiencyHamaguchi et al. (2026). Laboratory studies utilizing gnotobiotic mouse models have proven that mice lacking the sulfatase enzyme do not develop bacterial constipation, highlighting how this cooperative bacterial degradation is a primary driver of the condition.
Without this slippery mucin barrier, the walls of the canal network suffer from increased physical friction and elevated intestinal permeability. This thinning of the mucus allows microscopic particles to leak across the intestinal lining, triggering mild inflammatory responses that further disrupt normal biological operationsHamaguchi et al. (2026). Consequently, no matter how much water is introduced into the system, the lack of fundamental bacterial lubrication ensures that the transported material remains dry, hard, and completely immobile.
How do beneficial bacteria actively regulate intestinal movement?
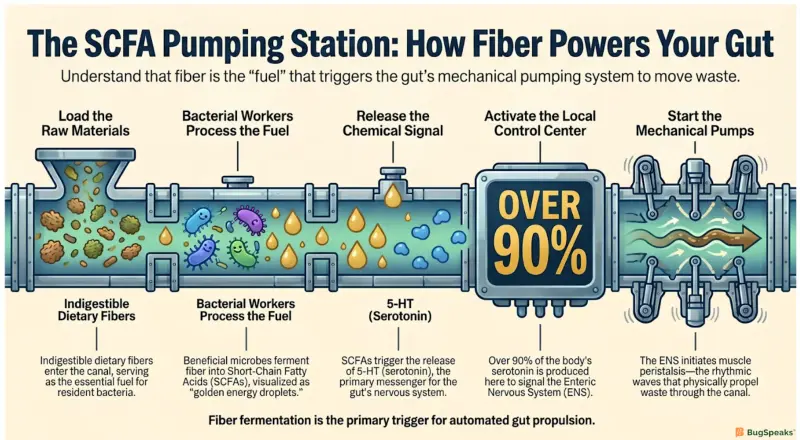
Beneficial microbes actively regulate intestinal movement by fermenting dietary fibers into specific chemical signals that instruct the gut's localized nervous system to contract and propel waste forward. In our canal network, passive lubrication is only one part of the equation; the system also requires active, rhythmic pumping mechanisms to keep the transported material moving efficiently. The flow-maintenance regulators drive this active propulsion by producing system-conditioning compounds scientifically known as Short-Chain Fatty Acids (SCFAs). When humans consume indigestible dietary fibers, beneficial bacteria such as Lactobacillus and Bifidobacterium ferment these fibers into primary SCFAs, including acetate, propionate, and butyrateBai et al. (2025).
These system-conditioning compounds serve multiple fundamental biological roles within the digestive tract. Butyrate, for example, acts as the primary energy source for the epithelial cells lining the colon, fulfilling up to seventy percent of their daily metabolic energy requirementsYu et al. (2026). By keeping the cellular infrastructure of the canal network energized and healthy, butyrate ensures that the physical walls can perform the synchronized muscle contractions required for proper transit. Furthermore, SCFAs function as direct chemical messengers that bind to specialized receptors, specifically G protein-coupled receptors, located on the surface of the intestinal liningBai et al. (2025).
Once these sensory receptors are activated by the system-conditioning compounds, they trigger specialized enterochromaffin cells to release 5-Hydroxytryptamine (5-HT), which is more commonly known as serotoninLi et al. (2025). Remarkably, the human gastrointestinal tract produces over ninety percent of the body’s total 5-HTBai et al. (2025). This powerful neurotransmitter directly stimulates the Enteric Nervous System (ENS), the localized brain governing the gut to initiate peristalsis, the rhythmic muscular waves that propel the transported material through the canal networkYu et al. (2026). The process is further assisted by Interstitial Cells of Cajal (ICC), which act as the electrical pacemakers for smooth muscle contractionBai et al. (2025).
When the gut microbial ecosystem lacks sufficient flow-maintenance regulators, the production of SCFAs drops drastically. The absence of these essential system-conditioning compounds means that the ENS never receives the 5-HT signaling required to trigger muscle contractions. The resulting loss of propulsive electrical force from the ICC leads directly to reduced flow efficiency, causing the transported material to stagnate. Restoring these specific fiber-fermenting bacteria is therefore essential to restarting the canal's automated propulsion systems.
What role do bile acids play in maintaining digestive transit?
Gut microbes transform primary liver secretions into secondary bile acids, which act as chemical keys to unlock specific receptors that strongly stimulate intestinal muscle contractions and fluid secretion. Beyond fermenting fiber, flow-maintenance regulators also manipulate powerful digestive fluids to keep the canal network operating at peak capacity. The human liver naturally secretes primary bile acids, such as cholic acid and chenodeoxycholic acid, which travel down into the digestive tract to assist in breaking down dietary fatsWen et al. (2026). However, a portion of these primary acids escapes early absorption in the small intestine and continues down into the deeper sections of the canal network.
Upon arriving in the colon, specific groups of flow-maintenance regulators intercept these primary digestive fluids. Using highly specialized bacterial enzymes, including bile salt hydrolases, these microbes chemically convert the primary bile acids into secondary bile acids, such as deoxycholic acid and lithocholic acidWen et al. (2026). This bacterial conversion is a vital physiological step because secondary bile acids possess a unique molecular shape that allows them to function as high-priority activation keys for the canal network's propulsion systems.
These secondary bile acids bind aggressively to a specific biological sensor called Takeda G Protein-Coupled Receptor 5 (TGR5), which is embedded in the walls of the intestineBai et al. (2025). Activation of TGR5, alongside the Farnesoid X Receptor (FXR), triggers a rapid and powerful response: it stimulates the release of both 5-HT and Glucagon-like peptide-1 (GLP-1)Wen et al. (2026). Together, these neurochemical signals instruct the smooth muscles of the canal network to contract forcefully while simultaneously drawing water into the canal to soften the transported material.
In patients suffering from severely reduced flow efficiency, the gut microbiome often exhibits a dangerous imbalance. The specific flow-maintenance regulators responsible for creating secondary bile acids are depleted, leaving an excess of primary bile acids and a total failure to activate the TGR5 and FXR sensorsWen et al. (2026). Without the crucial secondary bile acid signals, the canal network loses its mechanical rhythm, leading to sluggish transit times and hardened transported material. Medical interventions, such as fecal microbiota transplantation, can successfully restore this bile acid conversion process, effectively turning the canal's propulsion engines back onWen et al. (2026).
How does a slow digestive system affect brain health and mood?
A delayed digestive system allows inflammatory markers to accumulate and alter neurotransmitter production, which sends distress signals to the brain and actively contributes to mood disorders like depression. The human body operates on an interconnected grid known as the Microbiota-Gut-Brain Axis (MGBA). When the canal network suffers from prolonged reduced flow efficiency, the stagnation of transported material has consequences that reach far beyond the abdomenYu et al. (2026). A blocked canal eventually causes physical degradation of the natural flow coating, allowing harmful bacterial byproducts, such as lipopolysaccharides, to leak into the surrounding tissues and the bloodstreamLi et al. (2025).
This toxic leakage triggers an immediate systemic alarm, causing the body’s immune system to deploy pro-inflammatory cytokines, specifically Interleukin-6 (IL-6) and Tumor Necrosis Factor-alpha (TNF-α)Yu et al. (2026). These inflammatory molecules act as chemical distress signals that travel up the vagus nerve, the primary neural communication cable linking the canal network directly to the brain. Once these inflammatory markers cross into the central nervous system, they directly alter brain chemistry, initiating neuroinflammation and significantly increasing the risk of depression and chronic anxietyYu et al. (2026).
Furthermore, this persistent intestinal inflammation hijacks the body's normal chemical manufacturing lines. Specifically, it disrupts the metabolism of tryptophan, a vital dietary amino acid. Under healthy conditions, tryptophan is used to synthesize 5-HT to stabilize mood and maintain gut motility. However, when the canal network is inflamed, enzymes redirect tryptophan away from 5-HT production and instead force it down the kynurenine pathway, generating neurotoxic compounds that damage brain cellsYu et al. (2026). This also reduces the levels of Brain-Derived Neurotrophic Factor (BDNF), a protein crucial for neuronal survival.
This molecular hijacking creates a devastating biological loop. The brain experiences a severe shortage of serotonin, leading to clinical depression, while the canal network simultaneously loses the localized serotonin required to maintain muscle contractionsYu et al. (2026). Additionally, the psychological stress response activates the Hypothalamic-Pituitary-Adrenal (HPA) axis, flooding the body with cortisol, which further paralyzes the gut muscles and worsens the reduced flow efficiency. Correcting the microbial balance is therefore not just a digestive necessity, but a highly critical psychiatric intervention.
How can targeted probiotic therapies restore healthy intestinal flow?
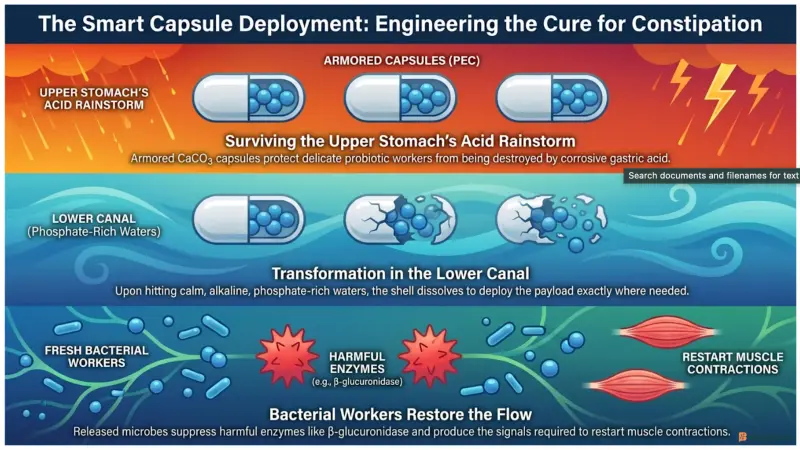
Targeted probiotic therapies restore healthy transit by utilizing smart delivery mechanisms to deploy beneficial microbes directly to the lower intestine, repairing the mucosal lining and reducing local inflammation. To permanently resolve reduced flow efficiency, scientists must reintroduce robust flow-maintenance regulators to the ecosystem. However, a major biological obstacle exists: the stomach is filled with highly corrosive gastric acid designed to destroy invading bacteria. If we simply swallow unshielded probiotics, the vast majority perish long before they reach the lower canal networkJeong et al. (2026).
To solve this, researchers have developed advanced encapsulation platforms, such as the Probiotics Encapsulated with Calcium Carbonate (PEC) system. By wrapping specific bacterial strains in a single, resilient layer of Calcium Carbonate (CaCO3), the microbes are effectively shielded against the harsh acidic environments of the upper digestive tractJeong et al. (2026). Once this armored payload reaches the lower canal network, the local alkaline environment and the presence of phosphate ions trigger a rapid structural transformation. The CaCO3 shell dissolves, deploying the live flow-maintenance regulators exactly where they are neededJeong et al. (2026).
Once successfully deployed, these targeted probiotic strains, such as Lacticaseibacillus rhamnosus and Bifidobacterium breve, go to work immediately. Clinical and animal models demonstrate that these newly introduced flow-maintenance regulators rapidly increase the water content of the transported material and significantly accelerate the intestinal transit rateJeong et al. (2026). They also proactively inhibit the activity of dangerous microbial enzymes like β-glucuronidase and nitroreductase, which are known to produce intestinal toxins, degrade the natural flow coating, and exacerbate tissue inflammationJeong et al. (2026).
Ultimately, this smart delivery approach results in a profound healing effect across the entire biological system. By safely delivering a multi-strain payload, the therapy suppresses systemic inflammatory markers like TNF-α and IL-6, tightens the cellular junctions of the intestinal walls, and continuously generates the system-conditioning compounds needed for daily operationLi et al. (2025). The canal network regains its natural flow coating and automated rhythmic pumping, scientifically proving that the secret to curing constipation lies in precision microbial engineering rather than merely increasing daily water intake.
Visualize the process- https://youtu.be/Pr6Kj5Nu2OE
Reference
Hamaguchi, T., Gibo, N., Ohara, M., Ito, M., Ogura, T., Takeda, J. I., … Ohno, K. (2026). Bacterial constipation: Mucin-degrading intestinal commensal bacteria cause constipation. Gut Microbes, 18(1). https://doi.org/10.1080/19490976.2025.2596809
Yu B, Zhao W, Tao L and Li K (2026) The microbiota-gut-brain axis perspective: mechanisms and intervention strategies for the comorbidity of chronic constipation and depression. Front. Microbiol. 17:1800520. doi: 10.3389/fmicb.2026.1800520
Li E, Wang J, Guo B and Zhang W (2025) Effects of short-chain fatty acid-producing probiotic metabolites on symptom relief and intestinal barrier function in patients with irritable bowel syndrome: a double-blind, randomized controlled trial. Front. Cell. Infect. Microbiol. 15:1616066. doi: 10.3389/fcimb.2025.1616066
Wen D, Liu S, Wu Y, Zhang H and Zhang K (2026) Fecal microbiota transplantation improves functional constipation through the gut microbiome–bile acid–receptor axis. Front. Med. 13:1751593. doi: 10.3389/fmed.2026.1751593
Jeong, S. N., & Kim, M. J. (2026). Smart CaCO3-coated probiotics relieve constipation by enhancing intestinal motility and modulating microbial enzymes in a loperamide-induced rat model. Scientific reports, 16(1), 7040. https://doi.org/10.1038/s41598-026-37864-6
Bai, X., Ihara, E., Tanaka, Y., Minoda, Y., Wada, M., Hata, Y., Esaki, M., Ogino, H., Chinen, T., & Ogawa, Y. (2025). The interplay of gut microbiota and intestinal motility in gastrointestinal function. Journal of smooth muscle research = Nihon Heikatsukin Gakkai kikanshi, 61, 51–58. https://doi.org/10.1540/jsmr.61.51